
Tot conținutul iLive este revizuit din punct de vedere medical sau verificat pentru a vă asigura cât mai multă precizie de fapt.
Avem linii directoare de aprovizionare stricte și legătura numai cu site-uri cu reputație media, instituții de cercetare academică și, ori de câte ori este posibil, studii medicale revizuite de experți. Rețineți că numerele din paranteze ([1], [2], etc.) sunt link-uri clickabile la aceste studii.
Dacă considerați că oricare dintre conținuturile noastre este inexactă, depășită sau îndoielnică, selectați-o și apăsați pe Ctrl + Enter.
Ovar
Expert medical al articolului

Ultima examinare: 04.07.2025

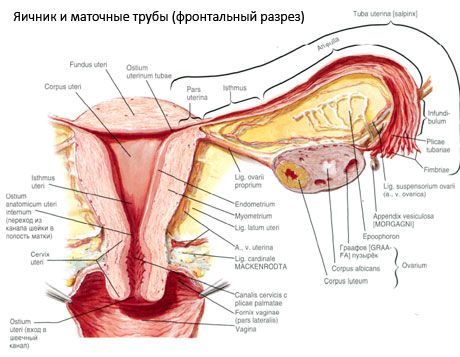
Ovarul (ovarium; grecesc oophoron) este un organ pereche, o glandă reproducătoare feminină, situată în cavitatea pelviană, în spatele ligamentului lat al uterului. În ovare se dezvoltă și se maturizează celulele reproducătoare feminine (ovulele), iar hormonii sexuali feminini intră în sânge și limfă. Ovarul are o formă ovoidă, oarecum aplatizată în direcție anteroposterioră. Culoarea ovarului este rozalie. Pe suprafața ovarului unei femei care a născut sunt vizibile depresiuni și cicatrici - urme de ovulație și transformarea corpului lutea. Ovarul cântărește 5-8 g. Dimensiunile ovarului sunt: lungimea 2,5-5,5 cm, lățimea 1,5-3,0 cm, grosimea - până la 2 cm. Ovarul are două suprafețe libere: suprafața medială (facies medialis), orientată spre cavitatea pelviană, acoperită parțial de trompele uterine, și suprafața laterală (facies lateralis), adiacentă peretelui lateral al pelvisului, până la o depresiune ușor pronunțată - fosa ovariană. Această fosă este situată în unghiul dintre vasele iliace externe acoperite cu peritoneu în partea superioară și arterele uterine și obturatoare în partea inferioară. În spatele ovarului, ureterul părții corespunzătoare trece retroperitoneal de sus în jos.
Suprafețele ovarului trec într-o margine liberă (posterioară) convexă (margo liber), în față - în marginea mezenterică (margo mesovaricus), atașată prin intermediul unui pli scurt al peritoneului (mezenterul ovarului) de foița posterioară a ligamentului lat al uterului. Pe această margine anterioară a organului există o depresiune canelată - hilul ovarului (hilum ovarii), prin care artera și nervii intră în ovar, iar venele și vasele limfatice ies. Ovarul are, de asemenea, două capete: capătul tubular superior rotunjit (extremitas tubaria), orientat spre trompele uterine, și capătul uterin inferior (extremitas utenna), conectat la uter prin ligamentul ovarian (lig. ovarii proprium). Acest ligament, sub forma unui cordon rotund de aproximativ 6 mm grosime, merge de la capătul uterin al ovarului până la unghiul lateral al uterului, situat între cele două foițe ale ligamentului lat al uterului. Aparatul ligamentos al ovarului include și ligamentul suspensor al ovarului (lig.suspensorium ovarii), care este un pliu al peritoneului ce pornește de la peretele pelvisului mic până la ovar și conține vasele ovariene și fascicule de fibre fibroase în interior. Ovarul este fixat de un mezenter scurt (mesovarium), care este o duplicație a peritoneului ce pornește de la foița posterioară a ligamentului lat al uterului până la marginea mezenterică a ovarului. Ovarele în sine nu sunt acoperite de peritoneu. Cea mai mare franjă ovariană a trompelor uterine este atașată de capătul tubular al ovarului. Topografia ovarului depinde de poziția uterului, de dimensiunea acestuia (în timpul sarcinii). Ovarele sunt organe foarte mobile ale cavității pelvine mici.
 [
[ Vasele și nervii ovarului
Alimentarea cu sânge a ovarelor este asigurată de aa. et vv. ovariae et uterinae. Ambele artere ovariene (aa. ovaricae dextra et sinistra) își au originea pe suprafața anterioară a aortei, chiar sub arterele renale; cea dreaptă își are originea de obicei în aortă, iar cea stângă în artera renală. Îndreptându-se în jos și lateral de-a lungul suprafeței anterioare a mușchiului psoas major, fiecare arteră ovariană traversează ureterul în față (dând ramuri către acesta), vasele iliace externe, linia de delimitare și intră în cavitatea pelviană, fiind situată aici în ligamentul suspensor al ovarului. Urmând în direcția medială, artera ovariană trece printre frunzele ligamentului lat al uterului pe sub trompa uterină, dând ramuri către acesta, și apoi în mezenterul ovarului; intră în hilul ovarului.
Ramurile arterei ovariene se anastomozează larg cu ramurile ovariene ale arterei uterine. Scurgerea venoasă din ovare se realizează în principal în plexul venos ovarian, situat în regiunea hilului ovarian. De aici, fluxul sanguin trece în două direcții: prin vena uterină și vena ovariană. Vena ovariană dreaptă are valve și se varsă în vena cavă inferioară. Vena ovariană stângă se varsă în vena renală stângă, deși nu are valve.
Drenajul limfatic din ovare are loc prin vasele limfatice, în special abundent în zona porții organelor, unde se distinge plexul limfatic subovarian. Apoi, limfa este evacuată de-a lungul vaselor limfatice ovariene către ganglionii limfatici paraaortici.
Inervația ovarelor
Simpatic - asigurat de fibrele postganglionare din plexurile celiac (solar), mezenteric superior și hipogastric; parasimpatic - de nervii sacrali viscerali.
Structura ovarului
Suprafața ovarului este acoperită cu un epiteliu germinativ cu un singur strat. Sub acesta se află un înveliș dens de proteine din țesut conjunctiv (tunica albuginea). Țesutul conjunctiv al ovarului formează stroma sa (stroma ovarii), bogată în fibre elastice. Substanța ovarului, parenchimul său, este împărțită în straturi exterioare și interioare. Stratul interior, situat în centrul ovarului, mai aproape de poarta sa, se numește medulară (medulla ovarii). În acest strat, în țesutul conjunctiv lax, există numeroase vase de sânge și limfatice și nervi. Stratul exterior al ovarului - cortexul (cortex ovarii) este mai dens. Conține mult țesut conjunctiv, în care sunt localizați foliculii ovarieni primari (folliculi ovarici primarii), secundari (veziculari) (folliculi ovarici secundarii, s.vesiculosi), precum și foliculi maturi, foliculi graafieni (folliculi ovarici maturis), precum și corpi galbeni și atretici.
Fiecare folicul conține un ovul reproducător feminin, sau ovocit (ovocit). Ovulul are un diametru de până la 150 µm, este rotund, conține un nucleu, o cantitate mare de citoplasmă, care, pe lângă organitele celulare, conține incluziuni proteico-lipidice (gălbenuș), glicogen, necesar pentru nutriția ovulului. Ovulul își consumă de obicei rezerva de nutrienți în decurs de 12-24 de ore de la ovulație. Dacă fertilizarea nu are loc, ovulul moare.
Ovulul uman are două membrane care îl acoperă. În interior se află citolema, care este membrana citoplasmatică a ovulului. În exteriorul citolemei se află un strat de așa-numite celule foliculare, care protejează ovulul și îndeplinesc o funcție de formare a hormonilor - secretă estrogeni.
Poziția fiziologică a uterului, trompelor și ovarelor este asigurată de aparatele de suspendare, fixare și susținere care unesc peritoneul, ligamentele și țesutul pelvin. Aparatul de suspendare este reprezentat de formațiuni pereche, acesta include ligamentele rotunde și late ale uterului, ligamentele proprii și ligamentele suspendate ale ovarelor. Ligamentele late ale uterului, ligamentele proprii și suspensoare ale ovarelor mențin uterul în poziția mediană. Ligamentele rotunde trag fundul uterin înainte și îi asigură înclinarea fiziologică.
Aparatul de fixare (ancorare) asigură poziția uterului în centrul pelvisului subțire și face practic imposibilă deplasarea acestuia în lateral, înainte și înapoi. Însă, deoarece aparatul ligamentos se îndepărtează de uter în secțiunea sa inferioară, sunt posibile înclinări ale uterului în diferite direcții. Aparatul de fixare include ligamente situate în țesutul lax al pelvisului și care se extind de la secțiunea inferioară a uterului până la pereții laterali, anteriori și posteriori ai pelvisului: ligamentele sacromagice, cardinale, uterovezicale și vezicopubiene.
Pe lângă mezovariu, se disting următoarele ligamente ovariene:
- ligamentul suspensor al ovarului, denumit anterior ligament infundibulopelvic. Este un pliu al peritoneului prin care trec vase de sânge (a. et v. ovarica) și vase limfatice și nervi ai ovarului, întins între peretele lateral al pelvisului, fascia lombară (în zona de divizare a arterei iliace comune în externă și internă) și capătul superior (tubar) al ovarului;
- Ligamentul propriu al ovarului trece printre frunzele ligamentului uterin lat, mai aproape de frunza posterioară, și leagă capătul inferior al ovarului de marginea laterală a uterului. Ligamentul propriu al ovarului este atașat de uter între începutul trompei uterine și ligamentul rotund, în spatele și deasupra acestuia din urmă. În grosimea ligamentului trec ramurile rotunde ale trompei ovarii, care sunt ramurile terminale ale arterei uterine;
- ligamentul apendiculo-ovarian Ligamentul se întinde de la mezenterul apendicelui până la ovarul drept sau ligamentul lat al uterului sub forma unui pliu al peritoneului. Ligamentul este inconstant și se observă la 1/2 - 1/3 dintre femei.
Aparatul de susținere este reprezentat de mușchii și fascia planșeului pelvin, împărțit în straturi inferioare, medii și superioare (interioare).
Cel mai puternic este stratul muscular superior (interior), reprezentat de mușchiul pereche care ridică anusul. Acesta este format din fascicule musculare care se extind în evantai de la coccis la oasele pelvine în trei direcții (pubococcygeus, iliococcygeus și ischiococcygeus). Acest strat de mușchi este numit și diafragmă pelviană.
Stratul mijlociu de mușchi este situat între simfiză, oasele pubian și ischial. Stratul mijlociu de mușchi - diafragma urogenitală - ocupă jumătatea anterioară a orificiului pelvin, prin care trec uretra și vaginul. În secțiunea anterioară, între foițele sale, se află fascicule musculare care formează sfincterul extern al uretrei, iar în secțiunea posterioară se află fascicule musculare care merg în direcția transversală - mușchiul transvers profund al perineului.
Stratul inferior (exterior) al mușchilor planșeului pelvin este alcătuit din mușchi superficiali, a căror formă seamănă cu numărul 8. Aceștia includ sfincterul bulbocavernos, ischiocavernos, sfincterul anal extern și mușchiul perineal transvers superficial.
Ontogeneza ovarelor
Procesul de creștere și atrezie a foliculilor începe la 20 de săptămâni de sarcină, iar până la naștere, în ovarele fetei rămân până la 2 milioane de ovocite. Până la menarhă, numărul lor scade la 300 de mii. Pe întreaga perioadă a vieții reproductive, nu mai mult de 500 de foliculi ating maturitatea și ovulează. Creșterea inițială a foliculilor nu depinde de stimularea cu FSH, este limitată, iar atrezia apare rapid. Se crede că, în loc de hormoni steroizi, peptidele autocrine/paracrine locale sunt principalul regulator al creșterii și atreziei foliculilor primari. Se crede că procesul de creștere și atrezie a foliculilor nu este întrerupt de niciun proces fiziologic. Acest proces continuă la toate vârstele, inclusiv în perioada intrauterină și menopauză, și este întrerupt de sarcină, ovulație și anovulație. Mecanismul care declanșează creșterea foliculilor și numărul acestora în fiecare ciclu specific sunt încă neclar.
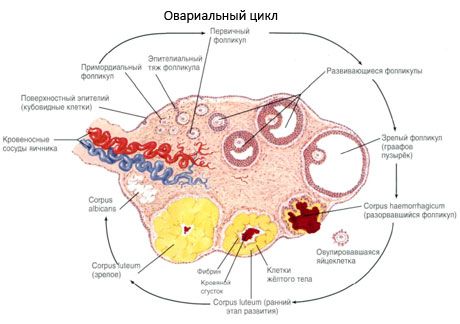
În timpul dezvoltării sale, foliculul trece prin mai multe etape de dezvoltare. Celulele germinale primordiale provin din endodermul sacului vitelin, alantoidă și migrează în zona genitală a embrionului la 5-6 săptămâni de sarcină. Ca urmare a diviziunii mitotice rapide, care continuă de la 6-8 săptămâni până la 16-20 săptămâni de sarcină, în ovarele embrionului se formează până la 6-7 milioane de ovocite, înconjurate de un strat subțire de celule granuloase.
Foliculul preantral - ovocitul este înconjurat de o membrană (Zona pellucida). Celulele granuloase care înconjoară ovocitul încep să prolifereze, creșterea lor depinde de gonadotropine și se corelează cu nivelul de estrogeni. Celulele granuloase sunt ținta FSH-ului. În stadiul de folicul preantral, celulele granuloase sunt capabile să sintetizeze trei clase de steroizi: induc predominant activitatea aromatazei, principala enzimă care transformă androgenii în estradiol. Se crede că estradiolul este capabil să crească numărul propriilor receptori, exercitând un efect mitogenic direct asupra celulelor granuloase, independent de FSH. Este considerat un factor paracrin care amplifică efectele FSH-ului, inclusiv activarea proceselor de aromatizare.
Receptorii FSH apar pe membranele celulelor granuloase imediat ce începe creșterea foliculului. O scădere sau o creștere a nivelului de FSH duce la o modificare a numărului de receptori ai acestuia. Această acțiune a FSH este modulată de factorii de creștere. FSH acționează prin intermediul proteinei G, sistemul adenilat ciclazei, deși steroidogeneza în folicul este reglată în principal de FSH, mulți factori sunt implicați în acest proces: canale ionice, receptori tirozin kinazici, sistemul fosfolipazic al mesagerilor secundari.
Rolul androgenilor în dezvoltarea timpurie a foliculilor este complex. Celulele granuloase au receptori androgenici. Aceștia nu sunt doar un substrat pentru aromatizarea indusă de FSH la estrogeni, ci pot, de asemenea, să amplifice aromatizarea la concentrații scăzute. Când nivelurile de androgeni cresc, celulele granuloase preantrale aleg preferențial nu calea de aromatizare la estrogeni, ci calea androgenică mai simplă prin 5a-reductază pentru a se converti într-un androgen care nu poate fi convertit în estrogen, inhibând astfel activitatea aromatazei. Acest proces inhibă, de asemenea, formarea receptorilor FSH și LH, oprind astfel dezvoltarea foliculilor.
În procesul de aromatizare, foliculul cu niveluri ridicate de androgeni trece prin procese de atrezie. Creșterea și dezvoltarea foliculului depind de capacitatea sa de a converti androgenii în estrogeni.
În prezența FSH, substanța dominantă a lichidului folicular va fi estrogenul. În absența FSH - androgenii. LH este în mod normal absent în lichidul folicular până la mijlocul ciclului. De îndată ce nivelul LH crește, activitatea mitotică a celulelor granuloase scade, apar modificări degenerative și nivelul androgenilor din folicul crește. Nivelul de steroizi din lichidul folicular este mai mare decât în plasmă și reflectă activitatea funcțională a celulelor ovariene: celulele granuloase și tecale. Dacă singura țintă pentru FSH sunt celulele granuloase, atunci LH are numeroase ținte - celulele tecale, celulele stromale și luteinice și celulele granuloase. Atât celulele granuloase, cât și cele tecale au capacitatea de steroidogeneză, dar activitatea aromatazei predomină în celulele granuloase.
Ca răspuns la LH, celulele tecale produc androgeni, care sunt apoi transformați de celulele granuloase în estrogeni prin aromatizare indusă de FSH.
Pe măsură ce foliculul crește, celulele tecale încep să exprime gene pentru receptorii LH, P450 sec și 3beta-hidroxisteroid dehidrogenaza, factorul de creștere asemănător insulinei (IGF-1) crește sinergic cu LH expresia genelor, dar nu stimulează steroidogeneza.
Steroidogeneza ovariană este întotdeauna dependentă de LH. Pe măsură ce foliculul crește, celulele tecale exprimă enzima P450c17, care formează androgeni din colesterol. Celulele granuloase nu au această enzimă și sunt dependente de celulele tecale pentru a produce estrogeni din androgeni. Spre deosebire de steroidogeneză, foliculogeneza este dependentă de FSH. Pe măsură ce foliculul crește și nivelurile de estrogen cresc, se activează un mecanism de feedback - producția de FSH este inhibată, ceea ce, la rândul său, duce la o scădere a activității aromatazei foliculului și, în cele din urmă, la atrezie foliculară prin apoptoză (moarte celulară programată).
Mecanismul de feedback al estrogenilor și FSH-ului inhibă dezvoltarea foliculilor care au început să crească, dar nu și a foliculului dominant. Foliculul dominant conține mai mulți receptori FSH, care susțin proliferarea celulelor granuloase și aromatizarea androgenilor în estrogeni. În plus, căile paracrine și autocrine acționează ca un coordonator important al dezvoltării foliculului antral.
Regulatorul autocrin/paracrin este format din peptide (inhibină, activină, folistatină), care sunt sintetizate de celulele granuloase ca răspuns la FSH și pătrund în lichidul folicular. Inhibina reduce secreția de FSH; activina stimulează eliberarea de FSH din glanda pituitară și amplifică acțiunea FSH în ovar; folistatina suprimă activitatea FSH, posibil prin legarea de activină. După ovulație și dezvoltarea corpului galben, inhibina este sub controlul LH.
Creșterea și diferențierea celulelor ovariene sunt influențate de factorii de creștere asemănători insulinei (IGE). IGF-1 acționează asupra celulelor granuloase, provocând o creștere a adenozin monofosfatului ciclic (cAMP), progesteronului, oxitocinei, proteoglicanului și inhibinei.
IGF-1 acționează asupra celulelor tecale, provocând o producție crescută de androgeni. Celulele tecale, la rândul lor, produc factor de necroză tumorală (TNF) și factor de creștere epidermică (EGF), care sunt, de asemenea, reglate de FSH.
EGF stimulează proliferarea celulelor granuloase. IGF-2 este principalul factor de creștere din lichidul folicular, iar în acesta se găsesc și IGF-1, TNF-α, TNF-3 și EGF.
Perturbarea reglării paracrine și/sau autocrine a funcției ovariene pare să joace un rol în tulburările de ovulație și în dezvoltarea ovarelor polichistice.
Pe măsură ce foliculul antral crește, conținutul de estrogen din lichidul folicular crește. În punctul culminant al creșterii acestora, pe celulele granuloase apar receptori pentru LH, are loc luteinizarea celulelor granuloase și producția de progesteron crește. Astfel, în perioada preovulatorie, o creștere a producției de estrogen determină apariția receptorilor pentru LH, LH, la rândul său, provocând luteinizarea celulelor granuloase și producția de progesteron. O creștere a progesteronului reduce nivelul de estrogen, ceea ce aparent provoacă un al doilea vârf de FSH la mijlocul ciclului.
Se consideră că ovulația are loc la 10-12 ore după vârful LH și la 24-36 de ore după vârful estradiolului. Se consideră că LH stimulează diviziunea reductivă a ovocitului, luteinizarea celulelor granuloase și sinteza progesteronului și prostaglandinei în folicul.
Progesteronul amplifică activitatea enzimelor proteolitice, care împreună cu prostaglandinele participă la ruptura peretelui folicular. Vârful de FSH indus de progesteron permite ovocitului să iasă din folicul prin transformarea plasminogenului în enzima proteolitică plasmină și asigură un număr suficient de receptori LH pentru dezvoltarea normală a fazei luteale.
În decurs de 3 zile de la ovulație, celulele granuloase cresc în dimensiune și în ele apar vacuole caracteristice umplute cu pigment, luteină. Celulele teco-luteale se diferențiază de tecă și stromă și devin parte a corpului luteal. Capilarele care penetrează corpul luteal se dezvoltă foarte rapid sub influența factorilor de angiogeneză, iar odată cu îmbunătățirea vascularizației, producția de progesteron și estrogeni crește. Activitatea steroidogenezei și durata de viață a corpului luteal sunt determinate de nivelul de LH. Corpusul luteal nu este o formațiune celulară omogenă. Pe lângă 2 tipuri de celule luteale, acesta conține celule endoteliale, macrofage, fibroblaste etc. Celulele luteale mari produc peptide (relaxină, oxitocină) și sunt mai active în steroidogeneză, cu o activitate aromatazică mai mare și o sinteză mai mare de progesteron decât celulele mici.
Vârful progesteronului apare în a 8-a zi după vârful LH. Se observă că progesteronul și estradiolul sunt secretate episodic în faza luteală, în corelație cu producția pulsatilă de LH. Odată cu formarea corpului luteal, controlul producției de inhibină trece de la FSH la LH. Inhibina crește odată cu creșterea estradiolului înainte de vârful LH și continuă să crească după vârful LH, deși nivelurile de estrogen scad. Deși inhibina și estradiolul sunt secretate de celulele granuloase, acestea sunt reglate prin căi diferite. Scăderea inhibinei la sfârșitul fazei luteale contribuie la creșterea FSH pentru ciclul următor.
Corpusul galben scade foarte repede - în ziua a 9-a-11-a după ovulație.
Mecanismul degenerării este neclar și nu este legat de rolul luteolitic al estrogenilor sau de un mecanism legat de receptori, așa cum se observă în endometru. Există o altă explicație pentru rolul estrogenilor produși de corpul luteal. Se știe că estrogenii sunt necesari pentru sinteza receptorilor de progesteron în endometru. Estrogenii din faza luteală sunt probabil necesari pentru modificările legate de progesteron ale endometrului după ovulație. Dezvoltarea inadecvată a receptorilor de progesteron, ca o consecință a nivelurilor inadecvate de estrogen, este posibil un mecanism suplimentar pentru infertilitate și pierderi de sarcină timpurii, o altă formă de deficit de fază luteală. Se crede că durata de viață a corpului luteal se stabilește în momentul ovulației. Și cu siguranță va regresa dacă nu este susținută de gonadotropina corionică umană din cauza sarcinii. Astfel, regresia corpului luteal duce la o scădere a nivelurilor de estradiol, progesteron și inhibină. O scădere a inhibinei elimină efectul său supresor asupra FSH; O scădere a estradiolului și progesteronului permite o restaurare foarte rapidă a secreției de GnRH și eliminarea mecanismului de feedback de la glanda pituitară. O scădere a inhibinei și estradiolului, împreună cu o creștere a GnRH, dă naștere la FSH față de LH. O creștere a FSH duce la creșterea foliculului cu selecția ulterioară a unui folicul dominant, iar dacă sarcina nu are loc, începe un nou ciclu. Hormonii steroizi joacă un rol principal în biologia reproductivă și fiziologia generală. Aceștia determină fenotipul uman, afectează sistemul cardiovascular, metabolismul osos, pielea, bunăstarea generală și joacă un rol cheie în sarcină. Acțiunea hormonilor steroizi reflectă mecanismele intracelulare și genetice care sunt necesare pentru a transmite un semnal extracelular către nucleul celulei pentru a provoca un răspuns fiziologic.
Estrogenii penetrează difuz membrana celulară și se leagă de receptorii situați în nucleul celular. Complexul receptor-steroid se leagă apoi de ADN. În celulele țintă, aceste interacțiuni duc la exprimarea genelor, sinteza proteinelor și funcții specifice ale celulelor și țesuturilor.
Использованная литература